Proc. 56th Southern Pasture and Forage Crop Improvement Conference, Springdale, AR April 21-22, 2001
Germination and Dormancy of Eastern Gamagrass Seed
T. L. Springer tspringer@spa.ars.usda.gov, and C. L. Dewald
USDA, ARS, SPRRS
2000 18th Street
Woodward, OK 73801
and
G. E. Aiken
USDA, ARS, DBSFRC
6883 South State Hwy 23
Booneville, AR 72927
Introduction
Eastern gamagrass is a perennial, warm-season bunchgrass occurring in various habitats across the eastern and central USA, Mexico, Central America, northern South America, and the Caribbean Islands (Newell and de Wet, 1974; de Wet et al., 1982). It has been recognized as one of the most productive and palatable warm-season grasses of the southeastern USA (Dewald and Louthan, 1979) and for that reason has increased in popularity. Problems associated with stand establishment continue to limit the widespread use of eastern gamagrass.
An early concern related to seed quality was determining the pure seed unit, which is impossible to do visually. The pure seed unit is a caryopsis enclosed by a cupulate fruit case. Removal of ‘blank seed units’ will improve seed quality. Ahring and Frank (1968) successfully removed these seed units with air and floatation separation.
Germination of the caryopsis within the intact cupules is a seed quality concern. The caryopsis must imbibe moisture and the embryo must force open the cupule before germination is complete. Several chemical agents (potassium nitrate, sodium hypochlorite, gibberellic acid, carbon dioxide) have been used unsuccessfully to stimulate seed germination (Ahring and Frank, 1968; Anderson, 1985). Cold-moist stratification apparently weakens the cupule enough to allow the caryopsis to imbibe sufficient water to germinate. Ahring and Frank (1968) reported that stratification at 5 to 10 oC for 6 to 8 wk enhanced seed germination to 60 to 80 % of all viable seeds. Anderson (1985) reported similar findings except that germination percentages were lower for his seed lots. Ahring and Frank (1968) also found differences among seed lots for germination and response to prechill duration.
In contrast, naked caryopses, i.e., with the cupulate fruit case removed, germinate readily (Anderson, 1985). An alternative to seeding intact cupules would be to plant caryopses. An advantage of planting caryopses is that pastures could be seeded to known plant populations. The disadvantage to this is that the specialized equipment needed to hull gamagrass without significantly damaging the caryopsis has not yet been developed.
Complementing the above experiments, we have studied the physical biology of the cupulate fruit case and its role in dormancy of gamagrass. We have also determined: 1) the caryopsis weight distribution, 2) the effects of caryopsis weight on germination, 3) the role of the cupulate fruit case in seed dormancy, and 4) the force required to open the cupulate fruit case.
Materials and Methods
Seed of ‘Iuka’ and ‘Pete’ eastern gamagrass were separated into four samples each containing 1000-seed. Caryopses were extracted from the cupulate fruit case by hand and individually weighed and placed into five weight classes (1.0-10.0, >10.0-20.0, >20.0-30.0, >30.0-40.0, and >40.0 mg caryopsis-1). For each weight class, caryopses were weighed and counted. Mean caryopsis weight was calculated by dividing the total caryopses weight (mg) by the number of caryopses in each class. Caryopsis weight distribution percentages were calculated by dividing the number of caryopses in each class by the total number of caryopses in the sample.
Fifty caryopses of each weight class were germinated in plastic boxes on two layers of paper towel substrate moistened with 7 ml of distilled water. The germination test was conducted in a seed germinator set for alternating temperatures of 20 oC (dark) and 30 oC (light) for 16 and 8 h, respectively. Cumulative actual and abnormal germination counts were made at 7 and 14 d. At the end of 14 d the number of firm (non-germinated, apparently dormant) caryopses was recorded. Total potential germination was determined by adding the 14-d actual germination and the remaining firm caryopses. Seedling shoot and root lengths were used as a measure of seedling vigor. Five random seedlings were measured for each weight class at the 7-d germination count. The experimental design was a randomized block replicated four times and repeated twice. The caryopsis weight x cultivar interaction was used to test the heterogeneity of slopes between cultivars (Littell et al, 1991). Regression analysis was performed on germination data.
To determine the role of the cupulate fruit case on seed dormancy, an artificial fruit case was fashioned from an extruded plastic hexagonal tube and a caryopsis from the >30.0-40.0 weight class was placed inside it and paired with a naked caryopsis. The germination conditions were the same as in the previous germination experiment. Data for each cultivar were analyzed as a paired T-test with 48 pairs.
A 91.4-cm lever and a fulcrum placed 30.5 cm from the cupulate fruit case was used to determine the force required to open the fruit case after 0, 1, 2, 4, and 8 wk of prechill at 0 to 5 oC. A dissecting needle, attached to the short arm of the lever, was inserted 2 to 3 mm under the outer glume of the cupulate fruit case; at the other end of the lever hung a small bucket. Salt was added to the bucket until the fruit case hinge opened. The salt filled bucket was then weighted and the force calculated. The experiment consisted of four replications of 5 seed each at 0, 1, 2, 4, and 8 wk of prechill at 0 to 5 oC and repeated twice. Regression analysis was performed on force measurements.
Results and Discussion
Caryopses of both cultivars displayed unimodal weight distributions (Table 1), and the caryopsis made up approximately 32% of the weight of the intact fruit case. Eastern gamagrass seed matures continuously from mid-June through mid-August and the environmental conditions before, during, and after flowering significantly influence caryopsis weight. Interrupting the reproduction cycle (as by seed harvest) during grain filling to increases variation in caryopsis weight and also increases seed borne diseases.
Percentage of actual germination and total potential germination of weight-classed caryopses increased as caryopsis weight increased. A caryopsis weight x cultivar interaction did not occur suggesting homogeneity of slopes for cultivars (Littell et al., 1991). The relationship between percentage 7-d actual germination (Y) and milligram caryopsis weight (x) was Y = 51.48 – 344.99 x-1, r2 = 0.84 (Fig. 1); percentage 14-d actual germination (Y) and milligram caryopsis weight (x) was Y = 52.33 – 346.79 x-1, r2 = 0.83; and percentage total actual germination (Y) and milligram caryopsis weight (x) was Y = 60.28 – 403.79 x-1, r2 = 0.86 (Fig. 1). Differences in percentage firm (dormant) seed were attributed to caryopsis weight. The relationship between percentage 14-d firm seed (Y) and milligram caryopsis weight (x) was Y = 7.16 – 51.62 x-1, r2 = 0.35.
Caryopsis weight also influenced the length of seedling roots and shoots. Similar to caryopsis germination, no caryopsis weight x cultivar interaction was found. The relationship between centimeter root length (Y) and milligram caryopsis weight (x) was Y = -1.88 + 1.39 ln x, r2 = 0.75 (Fig. 2); and centimeter shoot length (Y) and milligram caryopsis weight (x) was Y = -0.08 + 0.47 ln x, r2 = 0.62 (Fig. 2).
The actual and potential caryopses germination in eastern gamagrass is significantly influenced by caryopsis weight. Similar findings have been reported for other native grass species (Kneebone and Cremer, 1955). Environmental conditions reducing plant stress during caryopsis filling may lead to an increase in photosynthates that enter the caryopsis resulting in a larger embryo and/or carbohydrate reserve. Larger caryopses tend to have more rapid and increased germination and are less susceptible to attack from microorganisms and produce larger, stronger seedlings more likely to compete successfully for moisture and light. The major cause of seedling death among prairie species is moisture stress (Blake, 1935).
The 7-d germination of caryopses banded with an artificial fruit case (Fig. 3) was significantly lower (P > 0.01) than the unbanded control. Unbanded caryopses of the cultivar ‘Iuka’ averaged 37.5 ± 7.1 % (mean ± SE) compared to 8.3 ± 4.0 % for the banded caryopses. Similarly, unbanded caryopses of ‘Pete’ averaged 50.0 ± 7.3 % compared to 4.2 ± 2.9 % for the banded caryopses. The percentage germination of unbanded caryopses for each cultivar was also in the range of germination percentages found for weight-classed caryopses.
Seed dormancy in these cultivars of eastern gamagrass is likely mechanical in nature. Coat-imposed dormancy may interfere with water uptake or gaseous exchange; the coat may contain chemical inhibitors or act as a barrier against the escape of inhibitors from the embryo; it may modify the light that reaches the embryo; or exert a mechanical restraint (Bewley and Black, 1982). In some species one or more of these actions may take place. Several grass species are known to exhibit coat-imposed dormancy, and one similar in nature to eastern gamagrass is that of buffalograss, Buchloe dactyloides (Nutt.) Engelm., (Ahring and Todd, 1977). Buffalograss caryopses, like eastern gamagrass, will germinate readily when removed from the fruit case.
Fruit case integrity is, in part, a function of moisture, chilling condition, and time. The relationship of the force in Newtons required to open a fruit case (Y) and the duration of prechill (x) was Y = 4.05 + 2.54 e–X, r2 = 0.41 (Fig. 4).
Under natural conditions, a caryopsis must first overcome the integrity of the cupulate fruit case to germinate. Given that the force required to open the fruit case does not change significantly after 4-wk of moist prechill, other factors, such as light, alternating temperatures, freezing and thawing, fire, rodents, and soil microorganisms, to name a few, may be involved in the slow decay of the fruit case. If the caryopsis remains viable over a long period of time and sufficient weathering to the fruit case occurs, then germination should proceed as normal. However, given a long decay process, chances are likely that the enclosed caryopsis will succumb to microorganisms. Thus, the previously reported benefits of prechill treatment are not due primarily to immediate effects on the cupule integrity, but may likely result from a predisposition of the cupule to more rapid deterioration from weathering processes.
Conclusions
The germination of ‘Pete’ and ‘Iuka’ eastern gamagrass seed was affected by caryopsis weight, in that, larger caryopses germinated better than smaller caryopses. This was not a surprise and is similar to many other plant species. Seed dormancy of these two cultivars was attributed to the indurate first glume and hard bony rachis internode which form the cupulate fruit case. The integrity of the tissue where the first glume and the rachis internode meet must be reduced before germination will proceed normally. Given that the force required to open the cupulate fruit case did not significantly change after a 4-wk moist prechill at 0 to 5 oC, late winter plantings would be adequate if soil temperatures and moisture were favorable. Planting prechill-conditioned seed in early spring would be an alternative if soil temperatures and moisture were unfavorable in late winter.
REFERENCES
Ahring, R. M., and H. Frank. 1968. Establishment of eastern gamagrass from seed and vegetative propagation. J. Range Manage. 21:27-30.
Ahring, R. M., and G. W. Todd. 1977. The bur enclosure of the caryopses of buffalograss as a factor affecting germination. Agron. J. 69:15-17.
Anderson, R. C. 1985. Aspects of the germination ecology and biomass production of eastern gamagrass (Tripsacum dactyloides L.). Bot. Gaz. 146:353-364.
Bewley, J. D., and M. Black. 1982. Physiology and biochemistry of seeds in relation to germination. II. viability, dormancy, and environmental control. Springer-Verlag. Berlin.
Blake, A. K. 1935. Viability and germination of seeds and early life history of prairie plants. Ecol. Monographs 5:405-460.
Dewald, C. L., and V. H. Louthan. 1979. Sequential development of shoot system components in eastern gamagrass. J. Range Manage. 32:147-151.
de Wet, J. M. J., J. R. Harlan, and D. Brink. 1982. Systematics of Tripsacum dactyloides (Gramineae). Am. J. Bot. 69:1251-1257.
Kneebone, W. R., and C. L. Cremer. 1955. The relationship of seed size to seedling vigor in some native grass species. Agronomy Journal 17:172-177.
Littell, R. C., R. J. Freund, and P. C. Philip. 1991. SAS system for linear models. 3rd ed., SAS Institute, Cary NC.
Newell, C. and J. M. J. de Wet. 1974. Morphological and cytological variability in Tripsacum dactyloides (Gramineae). Am. J. Bot. 61:652-664.
Table 1. Caryopsis weight and distribution percentages (based on caryopses numbers) of
Tripsacum dactyloides (cultivars ‘Iuka’ and ‘Pete’) by seed weight class.
|
Weight |
Weight ±SD |
Percent Distribution |
|
|
Iuka |
Pete |
||
|
– – – mg – – – |
– – – mg – – – |
– – – % – – – |
– – – % – – – |
|
1.0 – 10.0 |
7.12 ± 0.84 |
13.2 a† |
9.0 b |
|
> 10.0 – 20.0 |
15.78 ± 0.53 |
32.0 a |
26.5 a |
|
>20.0 – 30.0 |
26.05 ± 3.03 |
33.3 a |
41.0 a |
|
>30.0 – 40.0 |
32.94 ± 3.94 |
19.5 a |
18.6 a |
|
> 40.0 |
43.13 ± 0.51 |
2.0 a |
4.9 a |
|
Non-classed‡ |
22.17 ± 1.43 |
||
† Cultivar means within each seed weight class followed by the same letter are not
significantlydifferent at P # 0.05 [F-test].
‡ Average weight of non-classed caryopses of >1.0 mg.
| Fig. 1. Relationships of gamagrass caryopsis weight and actual germination at the end of a 14-d germination period (A) and caryopsis weight and total potential germination (14 d actual germination plus firm seed remaining after 14 d, B). Each data point is the mean ± SE of 8 experimental units. |
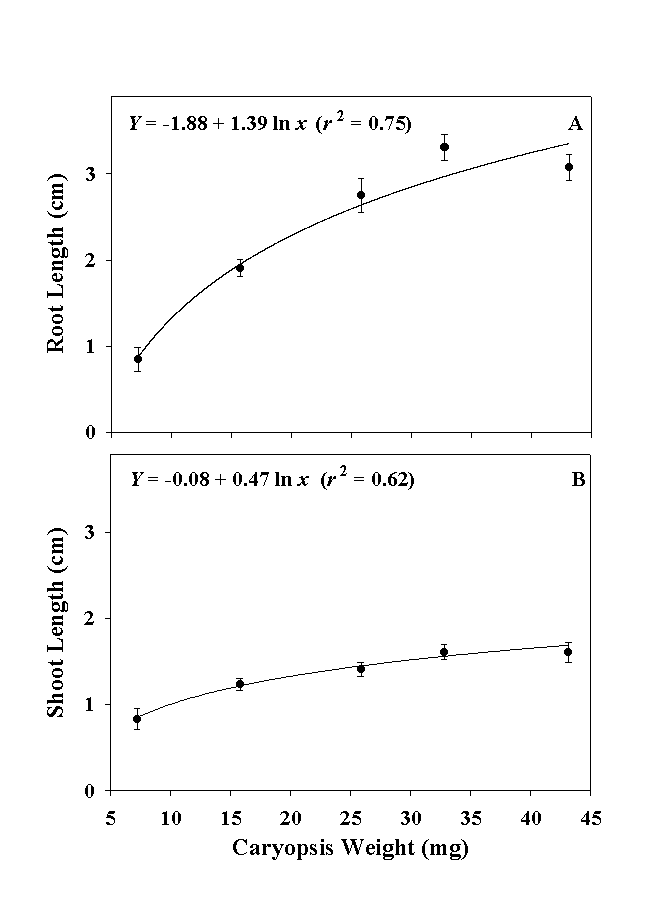 |
| Fig. 2. Relationships of gamagrass caryopsis weight and seedling root length (A) and caryopsis weight and seedling shoot length (B). Seedling measurements were made at end of the 7-d germination period. Each data point is the mean ± SE of 8 experimental units. |
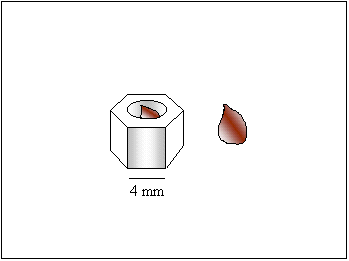 |
| Fig. 3. Drawing of artificial seed coat used to test for mechanical seed dormancy in gamagrass and a drawing of a naked caryopsis. The extruded hexagonal tube is 7 mm wide from face to face and 5 mm tall. Each of the six faces is 4 mm wide and 5 mm tall. The circular opening in the center of the hexagonal tube is 3 mm in diameter. |
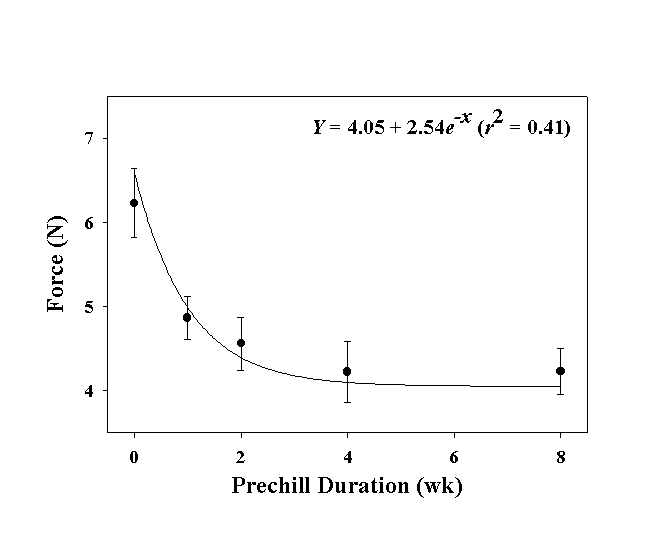 |
| Fig. 4. Relationship of duration of moist, prechill at 0-5 EC on the force required to open the cupulate fruit case of gamagrass. Each data point is the mean ± SE of 10 experimental units. |